转录因子是细菌中调控基因表达的关键蛋白质,它们能够识别并结合到DNA的特定序列上,从而启动或抑制基因的转录。细菌ChIP-seq可以帮助我们理解细菌中转录因子的调控网络,揭示其在细菌生长、代谢和适应环境等方面的作用。这种技术对于研究细菌的基因调控机制和适应性演化具有重要意义。本期我们带来了一篇来自中山大学医学院团队发表在Nature communications上的ChIP-seq文章,研究探讨了2',3'-环状鸟苷酸单磷酸(2',3'-cGMP)如何通过转录调节因子RSp0980调控青枯雷尔氏菌Ralstonia solanacearum的生物功能和毒力。

-
发表单位:中山大学医学院
-
发表期刊:Nature communications
-
发表日期:2023年11月10日
-
研究技术:ChIP-seq (His抗体)、RNA-seq、EMSA等
2023年11月10日,Nature communications杂志在线发表了中山大学医学院的研究论文“Regulation of the physiology and virulence of Ralstonia solanacearum by the second messenger 2′,3′-cyclic guanosine monophosphate”。该研究揭示了RSp0334的GGDEF结构域的不寻常功能以及细菌中2',3'-cGMP信号的特殊调节机制。爱基百客为该研究提供ChIP-seq的技术支持。

研究背景
此前研究发现双(3',5')-环二磷酸鸟苷单磷酸(bis-3',5'-c-di-GMP)在细菌中被广泛应用作为第二信使,用于调控细胞内的多种生物学功能。在该研究中,作者发现了2',3'-cGMP对青枯雷尔氏菌(Ralstonia solanacearum)中的重要生物功能、群体感知信号系统和毒力的调节作用,以及RSp0334和RSp0980在这一调节过程中的作用。这些发现为我们提供了对细菌中GGDEF结构域和2',3'-cGMP信号的特殊调节机制的深入了解。
研究内容
1. RSp0334在R. solanacearum中控制重要的生物功能
为了调查R. solanacearum中潜在的bis-3′,5′-c-di-GMP信号系统,研究对RpfR进行同源性搜索。选择了前9个同源物(按相似性排序)进一步研究,它们与RpfR的相似性在37.79%到42.63%之间。随后,研究构建了这9个同源物的缺失突变体。与R. solanacearum野生型菌株相比,RSp0254、RSp0334和RSp1155的缺失导致了运动活性分别降低了65.11%、70.22%和48%。此外,与野生型菌株相比,RSp0334突变株的生物膜形成减少了50.42%,而RSp0254和RSp1155突变株的生物膜形成分别增加了60.27%和39.17%。由于RSp0334的缺失破坏了R. solanacearum的生物膜形成和运动,因此选择其进行更深入的研究。
RSp0334的缺失对不同培养基中的细菌细胞生长几乎没有影响,但导致了运动、生物膜形成、纤维素酶产生和细胞外多糖(EPS)产生等表型的显著缺陷,而RSp0334的转录表达恢复了RSp0334缺失突变株的所有表型至野生型菌株的水平。
由于受测的表型受到吲哚丙酸信号和3-羟基棕榈酸甲酯(3-OH MAME)QS系统的控制,因此作者测试了RSp0334与这些信号系统之间是否存在关系。RT-qPCR和启动子-lacZ融合报告基因分析的结果显示,与R. solanacearum GMI1000野生型菌株相比,RSp0334突变株中phcB、solI和trpEG的表达水平显著降低。与这些结果一致的是,RSp0334突变株中3-OH MAME、N-辛酰-L-蛋氨酸内酯(C8-AHL)、N-癸酰-L-蛋氨酸内酯(C10-AHL)和吲哚丙酸的产量分别降低了78.13%、76.08%、72.74%和57.27%,而RSp0334的转录表达几乎完全恢复了这些信号的产量。
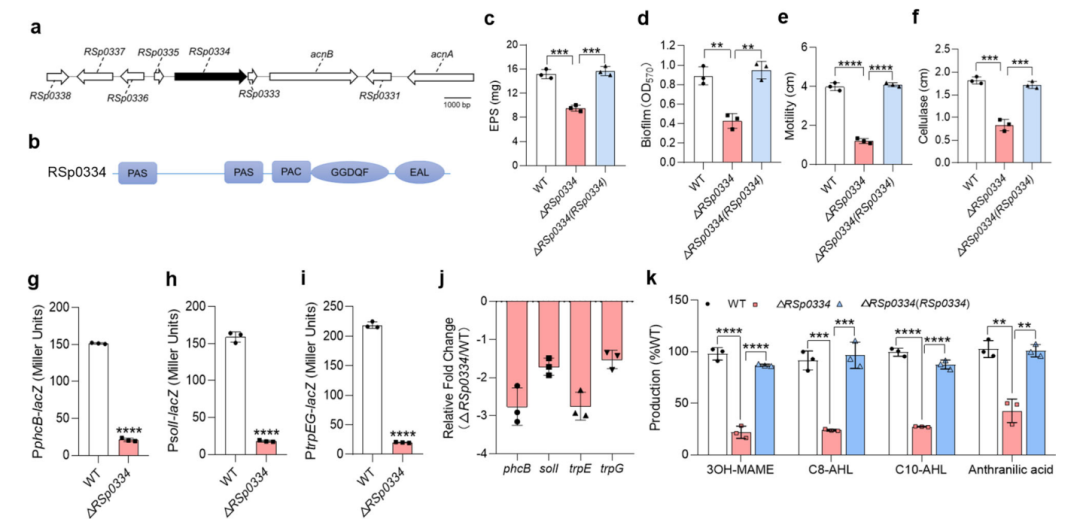
图1 RSp0334对毒力相关表型的影响。
2. RSp0334负调控R. solanacearum中的细胞内2′,3′-cGMP水平
由于RSp0334包含PAS-PAC-GGDQF-EAL结构域(图1b),作者继续研究其在环核苷酸分子代谢中的作用。HPLC分析显示,RSp0334的缺失导致细胞内未知分子水平显著增加,这种分子在转录表达中恢复到野生菌水平。由于这种分子的保留时间与标准的bis-3′,5′-c-di-GMP不同,因此从培养上清中分离和纯化了这种化合物。化合物的HR-ESI-MS分析显示该化合物可能具有C10H12N5O7P的分子式。最后,通过将NMR信息与2′,3′-cGMP进行比较,确认了从R. solanacearum中分离的化合物为2′,3′-cGMP,并且RSp0334突变株中的细胞内2′,3′-cGMP水平显著增加。
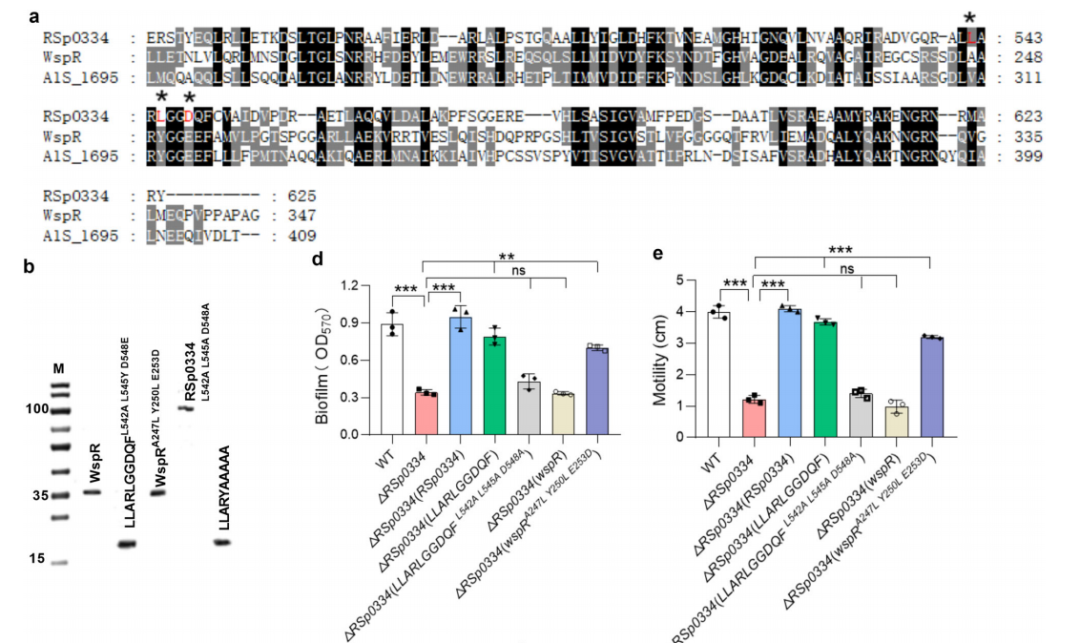
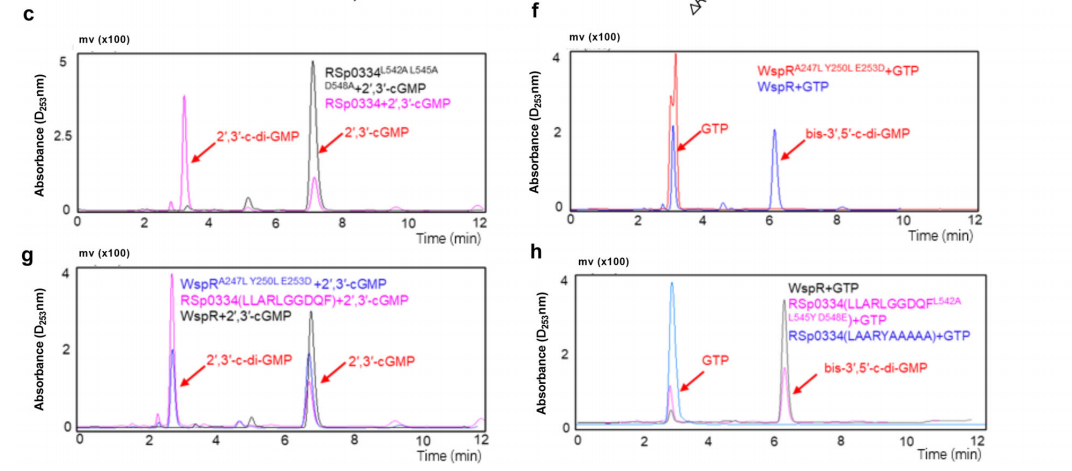
图2 RSp0334中关键酶活性位点的序列分析
3. RSp0334催化2′,3′-cGMP生成2′,3′-c-di-GMP
由于RSp0334负调控了R. solanacearum中的细胞内2′,3′-cGMP水平,作者假设它可能使用2′,3′-cGMP作为底物。为验证这一假设,通过亲和层析纯化了具有894个氨基酸和96 kDa计算分子量的RSp0334蛋白,并进行了体外酶活性实验。结果显示,在10分钟内,RSp0334蛋白催化了超过80%的2′,3′-cGMP,而该蛋白在体外不催化或降解2′,3′-cyclic adenosine monophosphate (2′,3′-cAMP)、GTP、bis-3′,5′-c-di-GMP或DNA。随后,作者纯化了产物,并通过HR-ESI-MS分析发现,产物的分子离子[M-H]-的m/z比为689.09003。在31P NMR谱图中,观察到产物具有两个化学位移(δ)为-1.20和-1.75的峰。这些结果表明,产物是一个环磷酸二鸟苷(cyclic di-GMP)。此外,在31P NMR谱图中存在两个明显的峰,表明产物是一个不对称的环磷酸二鸟苷,这一点也得到了1H NMR数据的支持。1H NMR谱图显示,化学位移(δ)为5.87、5.82、5.57、4.34、3.97、3.54和3.43的峰的积分值为1,这也证明了产物是不对称的,因为对称的bis-3′,5′-c-di-GMP应该具有均匀的积分值。因此得出结论,RSp0334将2′,3′-cGMP转化为2′,3′-c-di-GMP。
4. LLARLGGDQF基序催化2′,3′-cGMP生成2′,3′-c-di-GMP
由于RSp0334的GGDQF结构域催化2′,3′-cGMP生成2′,3′-c-di-GMP,而GGDEF结构域通常催化GTP生成bis-3′,5′-c-di-GMP,作者进一步研究了GGDQF结构域的详细功能。
为了测试Q549残基是否负责RSp0334的GGDQF结构域与bis-3′,5′-c-di-GMP合酶的GGDEF结构域之间的不同功能,生成了一个单点突变体RSp0334(GGDQF Q549E),其中残基Q549被谷氨酸(E)替换,将GGDQF结构域改变为GGDEF结构域。结果显示RSp0334(GGDQF Q549E)仍然表现出相同的催化2′,3′-cGMP的活性,但在将GTP转化为bis-3′,5′-c-di-GMP方面没有活性。
为了进一步探索GGDQF结构域的关键酶活性位点,作者生成了五种突变蛋白,突变了G546、G547、D548、Q549和F550这五个氨基酸残基。所有RSp0334(GGDQF)衍生物均通过亲和层析纯化,并用于体外酶活性实验。HPLC分析显示,只有Asp548的突变部分地减弱了RSp0334(GGDQF)的酶活性,而所有五个氨基酸残基的突变明显降低了RSp0334 GGDQF结构域的酶活性。RSp0334(GGDQF D548A)和RSp0334(AAAAA)的转录表达部分恢复了RSp0334缺失突变株的运动性和生物膜形成,使其恢复到野生型菌株的水平。
为了进一步确定RSp0334的酶活性是否需要其他氨基酸位点,作者与其他菌株的GGDEF结构域(GGEEF)的氨基酸进行序列比对。研究发现RSp0334具有Leu542和Leu545,这与两个bis-3′,5′-c-di-GMP DGCs中的残基不同。随后生成了突变蛋白RSp0334 L542A L545A D548A并测试了其酶活性。结果显示RSp0334 L542A L545A D548A对2′,3′-cGMP没有活性,并且RSp0334 L542A L545A D548A的转录表达不能恢复该突变体的生物膜形成和运动性,这表明L542、L545和D548三个氨基酸残基对RSp0334催化2′,3′-cGMP生成2′,3′-c-di-GMP的能力至关重要。
5. RSp0980是2′,3′-cGMP的受体,控制着RSp0334在R. solanacearum中调节的表型
作者在RSp0334缺失突变株中转录表达每个同源物,并测量转化菌株的运动性,发现只有RSp0980的转录表达才能恢复RSp0334突变株的运动性表型。因此,RSp0980被选定进行进一步研究。由于RSp0980的转录表达几乎完全恢复了RSp0334突变株的缺陷表型,继续研究RSp0980在调节重要的基础生物功能中的作用。作者构建了RSp0980的框内缺失突变,并发现RSp0980的缺失引起了与RSp0334突变株相同的表型变化,包括运动性、生物膜形成、EPS产生和纤维素酶产生,但不影响细菌在富营养或贫营养培养基中的生长速率。
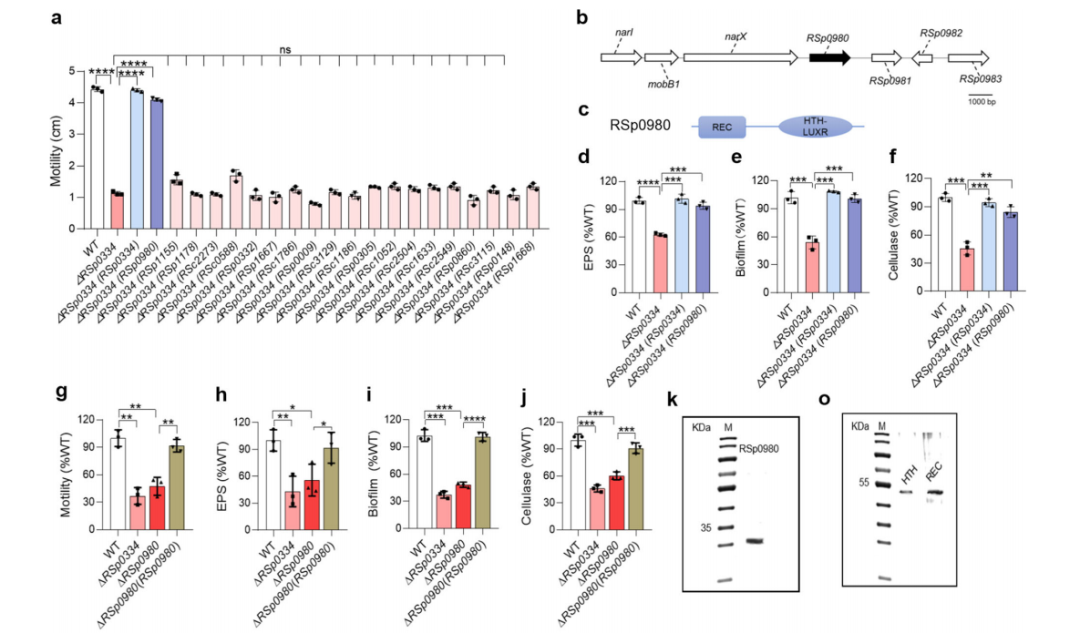
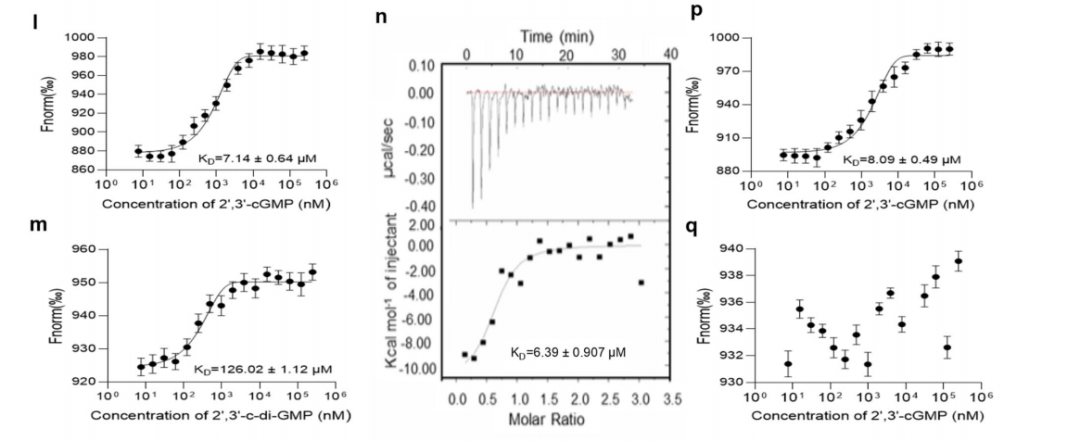
图3 RSp0980对RSp0334突变菌株的毒力相关表型的影响
6. RSp0980通过直接结合启动子区域来调节靶基因的表达
作者在RSp0980突变株中构建了PphcB-lacZ、PsolI-lacZ和PtrpEG-lacZ报告系统,分别对应合成3-OH MAME、AHL和蒽醌酸的基因。与ΔRSp0334突变株类似,RSp0980缺失导致phcB、solI和trpEG的表达水平降低(图4a-c)。因此,作者测量并比较了野生型、RSp0980突变株和回补菌株中3-OH MAME、C8-AHL、C10-AHL和蒽醌酸信号的产量。结果显示,RSp0980突变株的3-OH MAME、C8-AHL、C10-AHL和蒽醌酸的产量减少,而在转录RSp0980的情况下,信号产量几乎完全恢复正常水平。
RSp0980含有一个HTH结构域,预计与DNA结合密切相关。因此,作者通过电泳迁移率变化实验(EMSA)测试了RSp0980是否通过直接结合其启动子来实现对phcB、solI和trpEG的转录调节。实验结果显示phcB、solI和trpEG启动子DNA片段与RSp0980形成稳定的DNA-蛋白复合物,并且迁移速度比未结合的探针慢。与RSp0980突变株中EPS的产生一致,RSp0980通过直接结合其启动子来调节epsA的表达。此外,作者纯化了RSp0980的HTH结构域和REC结构域,并发现随着HTH结构域的增加,与标记的探针结合的量也增加,而在REC结构域和探针之间没有形成DNA-蛋白复合物,这表明HTH结构域结合到了靶基因的启动子上。
为了进一步确认RSp0980是RSp0334介导的信号系统的一个关键下游调节因子,并阐明其潜在的调节机制,作者采用ChIP-seq来鉴定RSp0980直接调控的靶基因。在潜在的靶基因中,phcB、solI、trpEG和epsA与R. solanacearum的生物功能和致病性相关。ChIP-seq数据显示RSp0980结合了多个基因启动子,这些启动子含有潜在的RSp0980结合位点,被确定为5’-GGAAATGGAC-3’。然后分别从phcB、solI、trpEG和epsA的启动子区域中删除了潜在的结合位点5’-CCTGCCCGAC-3’、5’-GCAAATTCCG-3’、5’-CGAACCCGAC-3’和5’-GGAAGTCGCC-3’。EMSA分析显示,当从探针中删除结合位点时,DNA-蛋白复合物没有形成,这表明这些特定片段对于RSp0980与启动子的结合是必不可少的。综上所述,这些结果表明RSp0980通过直接结合基因启动子中的特定区域来调节靶基因的表达。
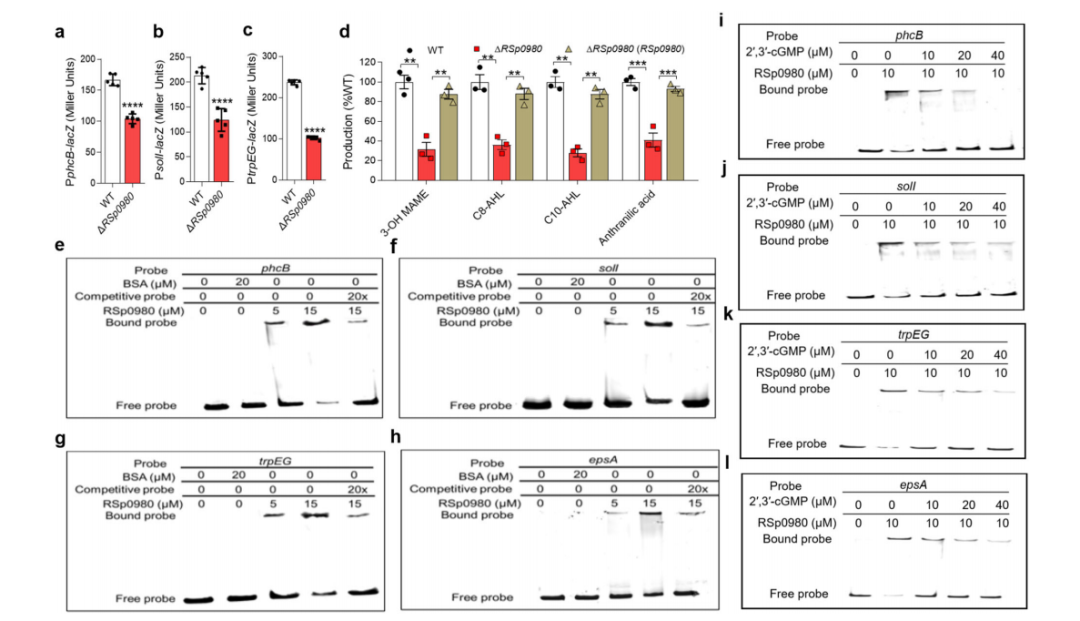
图4 RSp0980对QS信号系统的影响
7. 2',3'-cGMP消除了RSp0980与靶基因启动子DNA的结合
为了确定2',3'-cGMP与RSp0980的结合如何影响RSp0980的活性,作者通过EMSA检测了2',3'-cGMP对RSp0980与phcB、solI、trpEG和epsA启动子的结合效果。如图4所示,当混合物中存在2',3'-cGMP时,RSp0980与phcB、solI、trpEG和epsA启动子的结合受到抑制。随着2',3'-cGMP浓度的增加,与RSp0980结合的探针数量减少。与上述结果一致,外源2',3'-c-di-GMP或bis-(3',5')-c-di-GMP的添加并不影响RSp0980与epsA启动子的结合,而2',3'-cAMP在100μM时略微影响RSp0980与epsA启动子的结合,这表明RSp0980是2',3'-cGMP的特异效应物。
8. RSp0334/2',3'-cGMP/RSp0980信号系统控制着R. solanacearum的致病性和各种基因的表达
作者评估了RSp0334/2',3'-cGMP/RSp0980信号系统对R. solanacearum感染寄主植物的能力的影响。与被R. solanacearum GMI1000野生型菌株和回补菌株感染的植物相比,被RSp0334突变株和RSp0980突变株感染的植物出现了明显减少的萎蔫症状。接种21天后,接种野生型菌株、RSp0334补充菌株和RSp0980补充菌株的番茄植株的病情指数分别为4.0、3.8和3.8。RSp0334突变株和RSp0980突变株接种的植物的病情指数分别为3.0和2.7,分别比野生型菌株低25%和32.5%。
作者进一步量化了这些R. solanacearum菌株在番茄植株的根和茎中的菌落形成单位(CFU)值。接种后7天,R. solanacearum野生型菌株、RSp0334突变株、RSp0980突变株、RSp0334补充菌株和RSp0980补充菌株的CFU数分别为每克根组织2.45 × 109、9.32 × 105、2.21 × 107、5.18 × 108和4.12 × 108。在番茄茎上观察到类似的结果,这五种菌株在接种后7天的CFU数分别为每克茎组织9.13 × 109、1.93 × 106、1.09 × 107、1.51 × 109和1.16 × 109。这些结果表明,RSp0334/2',3'-cGMP/RSp0980信号系统在R. solanacearum的致病过程中发挥着重要作用。
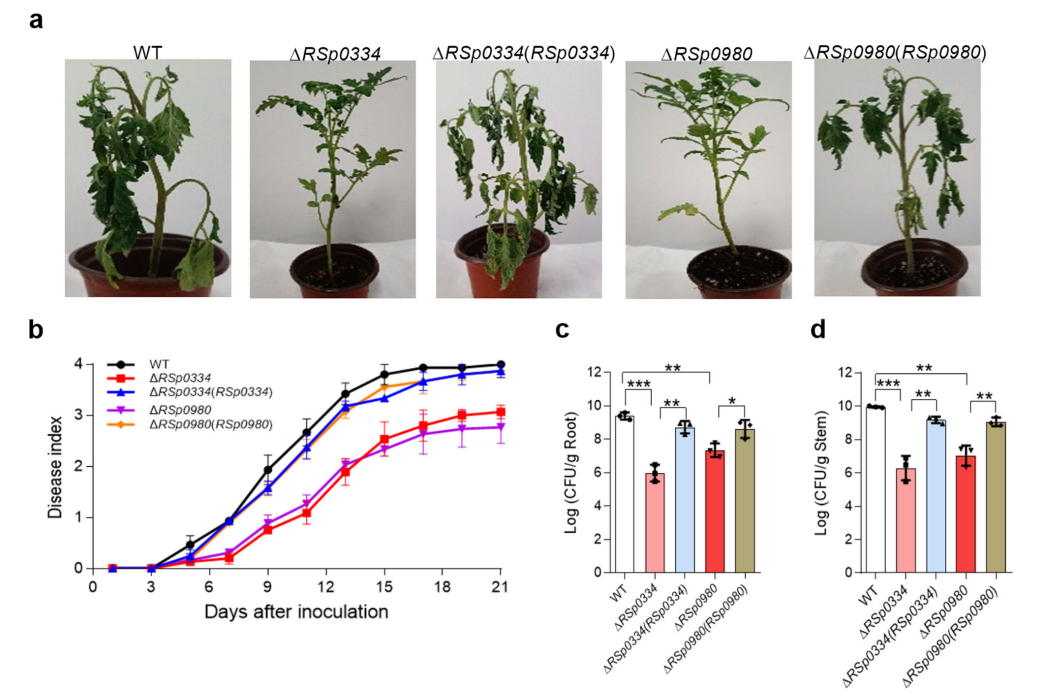
图5 RSp0334和RSp0980对番茄植株中R. solanacearum致病性的影响
小 结
研究报告了关于细菌中第二信使2',3'-环鸟苷酸单磷酸(2',3'-cGMP)的研究成果。2',3'-cGMP通过转录调节因子RSp0980控制青枯雷尔氏菌Ralstonia solanacearum中的重要生物功能、群体感知信号系统和毒力。此外,RSp0334的缺失导致细胞内2',3'-cGMP水平增加,从而改变了重要的表型。研究还发现,2',3'-cGMP、其受体以及具有LLARLGGDEF基序的进化的GGDEF结构域也存在于人类病原体沙门氏菌。这些发现揭示了RSp0334的GGDEF结构域的不寻常功能以及细菌中2',3'-cGMP信号的特殊调节机制。
爱基百客拥有10年表观遗传学研究服务的经验,我们提供CUT&Tag、ATAC-seq、ChIP-seq、DAP-seq等表观遗传学测序服务。服务范围涵盖实验设计、样本处理、数据分析到验证的全流程,旨在为客户提供全面、专业的技术支持。目前,我们在微生物的表观经验已相当丰富,辅助客户多次发表Nature Commun、Plant Commun、Nucleic Acids Res、New Phytol等国际高水平期刊,欢迎有需求的老师咨询~
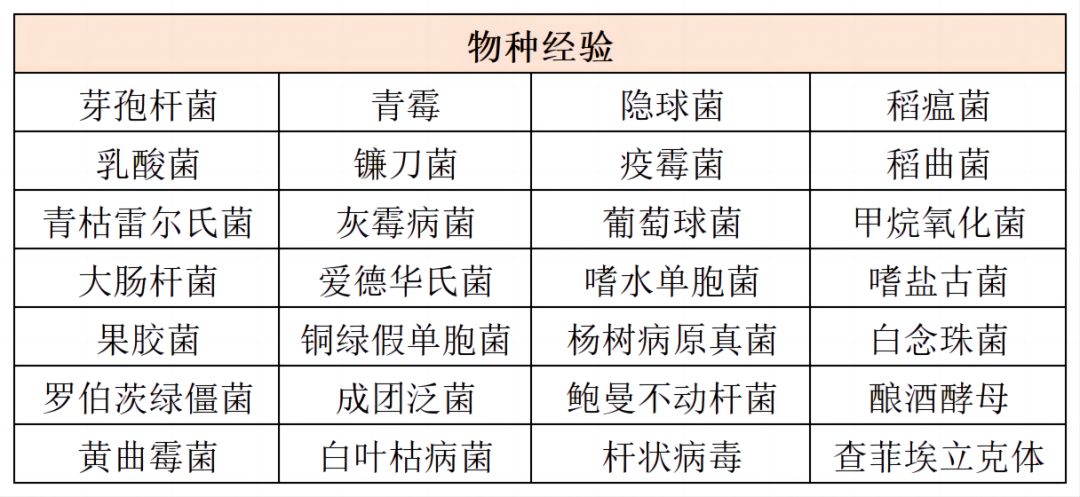
微生物表观经验部分汇总

· 爱基百客王牌产品ChIP-seq简介 ·
· ChIP-seq相关介绍 ·
ChIP-seq技术将染色质免疫共沉淀和二代测序技术结合,是研究体内蛋白质与DNA相互作用的有力工具,可用于组蛋白修饰、RNA聚合酶、转录因子和辅因子以及G4链体(G4)等方面的研究,技术成熟稳定。爱基百客ChIP-seq可提供:
-
ChIP-seq测序分析
Peak分析: Peak注释和分布分析,Peak关联基因的GO、KEGG的注释和富集分析, 转录因子和Motif分析等。
多样本差异分析:差异 Peak 分布情况统计,差异 Peak 关联基因GO、KEGG 功能注释与富集,转录因子预测,Motif 预测等。
· 后续验证
01 ChIP-qPCR
分析组蛋白修饰/转录因子与染色质区域的结合情况,揭示染色质状态和基因表达调控之间的关系,真实反映结合特性。
02 EMSA
基于DNA-蛋白质复合体在聚丙烯酰胺凝胶电泳(PAGE)中的迁移率不同,检测活化的与DNA结合的蛋白转录或调节因子。
03 双荧光素酶报告实验
检测转录因子与靶启动子的特异结合。
-
ChIP-seq+转录组关联分析
ChIP-seq和转录组关联分析可以做以下2个方面的研究:
1、DNA结合蛋白和基因表达调控:通过ChIP-seq技术可以确定DNA结合蛋白(如转录因子)的结合位点,然后与转录组数据结合分析,可以获得转录因子直接调控的靶基因,为全面理解转录因子调控功能提供依据。
2、组蛋白修饰和基因表达:ChIP-seq可以用于鉴定组蛋白修饰的位点,结合转录组数据可以了解这些修饰对基因表达的影响。
· 爱基百客ChIP-seq三大优势
优势一:项目经验丰富,研究物种200+种,累计实验2000余次。全面覆盖医口和农口等不同样本,不惧特殊样本(如脂肪组织、高淀粉组织和真菌类),抗体经验也极其丰富(多种组蛋白修饰、转录因子、标签抗体以及p300和RNApol II等均有涉及);
优势二:提供前期实验设计、测序、分析以及后期验证(ChIP-qPCR、EMSA)一站式服务;
优势三:项目文章多次发表于Cancer Cell、Nature Plants、Nature Metabolism以及Plant Cell等期刊。


![P1712 [NOI2016] 区间(线段树 + 贪心 + 双指针)](https://img-blog.csdnimg.cn/img_convert/4c30e02e3af7587428f7587fe6bc8127.png)